Mid-ocean spreading ridges – “the linked major mid-oceanic mountain systems of global extent” (IHO, 2008). Spreading ridges are distinguished from other ridges in this study (see definition of ridges). They were mapped by hand based on their appearance as ridge-like features that coincide with the youngest ocean crust as mapped by Müller et al. (1997) in their “EarthByte” digital age grid of the ocean floor. Spreading ridges that were not visible in the SRTM30_PLUS bathymetry (100 m contours; Becker et al., 2009) were not included in our interpretation, but there is otherwise no vertical size limitation on spreading ridges (they overlay the abyssal plains, hills or mountains classification layers in different locations). The mid-ocean spreading ridge covers the largest fraction of abyssal zone in the Arctic Ocean, where it characterises 4.76% of the area of abyssal zone, and it is absent from the Mediterranean and Black Sea. The greatest area of mid-ocean ridges occurs in the South Pacific Ocean where this feature type covers an area of 1,868,490 km2.
Mid-ocean ridges statistics (after Harris et al., 2014). The percentage areas refer to ocean basin areas and the percentage of the abyssal zone that is mid-ocean ridge.
| Ocean | Area km2 | Mid-ocean ridge area% | %Area of abyssal zone that is Mid-Ocean Ridge | Number of mid-ocean ridge segments | Average area of mid-ocean ridge segments km2 |
| Arctic Ocean | 254,630 | 1.96 | 4.76 | 5 | 50,930 |
| Indian Ocean | 1,547,910 | 2.17 | 2.46 | 34 | 45,530 |
| Mediterranean & Black Sea | 0 | 0 | 0 | 0 | 0 |
| North Atlantic | 677,630 | 1.51 | 2.01 | 5 | 135,530 |
| North Pacific | 840,300 | 1.02 | 1.22 | 24 | 35,010 |
| South Atlantic | 1,166,750 | 2.89 | 3.19 | 9 | 129,640 |
| South Pacific | 1,868,490 | 2.14 | 2.31 | 30 | 62,280 |
| Southern Ocean | 343,740 | 1.69 | 2.02 | 9 | 38,190 |
| All Oceans | 6,699,460 | 1.85 | 2.19 | 106 | 63,200 |
If all the water were removed from the ocean basins, there would be revealed a chain of mountains located in the middle of the oceans and encircling the world, like the seems of a baseball, along a total length of over 75,000 km. Mid-ocean ridges are created by the upwelling of basaltic lava and lateral rifting of ocean crust, thus forming a rift valley system. The mid-ocean ridges are the earth’s largest volcanic system, accounting for more than 75% of all volcanic activity on the planet. The heat from this volcanism is dispersed by hydrothermal circulation of seawater. Hot seawater venting from the seafloor supports strange benthic communities that have evolved to survive by using the hydrogen sulphide dissolved in the hot fluid.
The deepest parts of ocean basins are located in troughs or trenches, where ocean crust is being subducted beneath continental or other ocean crust into the earth’s mantle. Therefore, the deepest parts of the oceans are not located in the middle, but rather closer to land where the subduction trenches are found. The upwelling of lava, rifting apart of the mid-ocean ridges and the lateral spreading of ocean crust is the driving force of plate tectonics. The basaltic mid-ocean ridges are boundaries along which two ocean plates are pulled apart by the force of gravity. The pulling force is provided by the sinking of cooled, condensed ocean crust in deep ocean trenches at the other end of the ocean plate “conveyor belt”. Hence the mid-ocean ridge appears elevated because the surrounding ocean floor has cooled and subsided around it, which is different to mountains formed on land that are formed by land being pushed up by lateral compression. The age of the ocean crust is young nearest to the mid-ocean ridges and is generally older closer to the continents.
Deep sea vent communities
One of the most remarkable discoveries in oceanographic research is the existence of thriving colonies of animals located on the mid-ocean spreading ridges, feeding on sulfide-rich fluids escaping from hydrothermal vents. Deep sea vent communities comprise entirely separate ecosystems, decoupled from solar-powered life on the earth’s surface, having evolved to utilize organic matter synthesized by hydrogen sulfide reducing bacteria. Over 300 endemic species have been found near the vents, including corals, clams, shrimps, crabs and the now famous giant, red-tipped tubeworms, 4 m tall creatures that flourish in waters close to the hot springs.
Discovery
Hydrothermal vent communities were first discovered by marine geologists while investigating processes of geothermal heat flux from mid-ocean spreading ridges (van Dover, 2000). Heat derived from volcanic eruptions along the spreading ridges is released into the ocean, but in the 1960’s and early 1970’s the heat transfer mechanisms were not well understood. It was thought that seawater might move through the hot volcanic rocks to dissipate heat by convection processes, but hot springs had never been observed on the seafloor. Frequent earthquakes (80 per minute) and unusually warm water temperatures had been recorded near the ridges (Corliss et al., 1979), but there was no evidence for any deep sea “vents” as such.
On Feb. 17, 1977, the submersible Alvin, descended over 2,000 m to the Galapagos Spreading Center seeking evidence for hydrothermal circulation. A trail of white clamshells strewn across the dark volcanic rock background led the submersible onwards until, suddenly, the submersible crew observed “coming out of small cracks cutting across the lava terrain was warm shimmering water that quickly turned cloudy blue, as manganese and other chemicals in solution began to precipitate out of the warm water and were deposited on the lava surface, where they formed a brown stain.” (Robert Ballard, Oceanus Magazine, 1977). The first hydrothermal vent had been discovered.
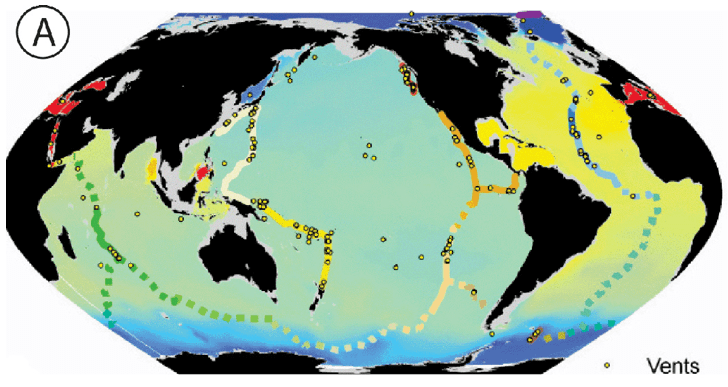
Hydrothermal vents: (A) Map showing vent sites and different ridge communities along the mid-ocean ridges of the world (from GOODS bioregionalisation). (B) Photograph taken by Charles Fisher, Woods Hole Oceanographic Institute of a vent-plume at the Mariner vent field on the Valu Fa Ridge near Fiji, South Pacific Ocean (C) giant tubeworms Riftia pachyptila at 13°N on the East Pacific Rise, Copyright Ifremer –Phare Expedition communication@ifremer.fr (D) swarming vent fauna, mid-Atlantic ridge (University of Bremen akluegel@uni-bremen.de).



Geothermal heat flux
The sulphides, sulphates and oxides precipitated from the hot (over 350°C) fluids escaping from deep sea vents form constructional, chimney-like features over 10 m in height. Vent chimneys are most commonly less than 20 m in height, but one over 45 m in height, named “Godzilla”, occurs on the Juan de Fuca Ridge. On exiting the chimney, the minerals in the hot fluid precipitate rapidly, forming what are described as black or white “smokers”. Mounds of precipitated pyrite-chalcopyrite several meters high cover the seafloor around the vents (Corliss et al., 1979).
The vents and their associated chemosynthetic communities are located on the axial ridge of the rift valley, where the geothermal heat flux is highest. Typically only a few km in width, axial ridges generally comprise a rocky substrate devoid of sediment cover, although ridge morphology varies depending upon the seafloor spreading rate: at slow spreading rates of less than 40 mm/yr, the paired flanks of the rift valley are far apart (5 to 15 km) and the valley depth is large (1 to 3 km deep). In contrast, fast spreading ridges do not exhibit a well-defined rift valley; rather the ridge supports a linear caldera or eruptive fissure 50 to 1000 m wide and only a few 10’s of meters deep (MacDonald et al., 1991). The east Pacific rise has the fastest spreading ocean crust, with rates of from 90 to 170 mm/yr.
Factors for conservation
Of significance for conservation is the correlation between the number of vent sites on slow versus fast spreading ridges; fast spreading ridges on the East Pacific Rise may have one active site for every 5 km of ridge crest, whereas vents along the slow spreading mid-Atlantic ridge occur only once in every 100 to 350 km (see van Dover, 2000, for details). Based on these figures, we can estimate that, whereas the fast-spreading ridges in the Pacific, Indian and Southern Oceans may support as many as 12,000 vent sites, the slow spreading mid-Atlantic ridge supports only on the order of 40 or so vents (i.e. there are perhaps only 40 hydrothermal vents along the entire length of the mid-Atlantic ridge in the North and South Atlantic Ocean). Clearly, the identification and conservation of the isolated mid-Atlantic ridge vent sites should be a priority for the global community, in terms of the protection of these ecosystems.
The communities that inhabit hydrothermal vents exhibit high degrees of endemism and diversity. The average biomass associated with vents is an order of magnitude at least larger than that associated with the surrounding deep sea environment. Clams and tubeworms characterise the assemblage of animals living near the vents. They host symbiotic, chemosynthetic bacteria that convert sulphides into organic matter. Other chemosynthetic organisms make filamentous mats and biofilms that provide food for grazers and deposit feeders. At the apex of the food web are predators and scavengers such as spider crabs, who move in from the surrounding abyssal areas. Some species of predators and scavengers have evolved to become vent specialists and are found only on vents (Van Dover, 2000).
Connectivity between hydrothermal vents
Factors that play a part in the connectivity between vents are the rate of vent formation, vent longevity, vent distribution, spacing between active vents, the temperature of venting seawater and bottom current regime at the vent site. The first factors are all related to the rate at which new ocean crust is formed. Thus a fundamental piece of information is the spreading rate of the ocean ridge system and the amount volcanic activity. As noted above, volcanic activity along the mid-Atlantic ridge is generally much less than in the Pacific, and the spacing between individual vent communities is much larger in the Atlantic than in the Pacific.
All of the water in the world ocean is estimated to flow through the mid-ocean ridges and escape through hydrothermal vents once in every 10 to 100 million years. For comparison, the rivers refill the oceans via the hydrological cycle about once every 30,000 years. Therefore the absolute flux of water via river inflow is equal to around 300 to 3,000 times the flux of seawater through hydrothermal vents. Although the total amount of water escaping from deep sea vents is not large, the fact that vents may occur along the entire length of the mid-ocean ridge, and also on the flanks of erupting undersea volcanos, suggests this process is of global significance.
Larvae spawning from a vent point source must rely upon bottom currents for dispersal, rather than the flow of water escaping from the vent. Axial ridges typically rise over 1,000 m in height above the level of the abyssal plains and thus interact with the flow of bottom water. Their linear geometry tends to deflect bottom currents to flow parallel to the ridge, enhancing larvae dispersal and colonization of vent sites along the axial ridge. In contrast, rift valleys and other basins perched between the parallel axial ridges may contain isolated volumes of water that have sluggish flow rates limiting larvae dispersal and colonization of vent sites. For these reasons the distribution patterns of today’s vent fauna display the strong imprint of the timing and geometry of ancient plate boundaries (Tunnicliffe and Fowler, 1996).
References
Becker, J.J., Sandwell, D.T., Smith, W.H.F., Braud, J., Binder, B., Depner, J., Fabre, D., Factor, J., Ingalls, S., Kim, S.H., Ladner, R., Marks, K., Nelson, S., Pharaoh, A., Trimmer, R., Von Rosenberg, J., Wallace, G., Weatherall, P., 2009. Global Bathymetry and Elevation Data at 30 Arc Seconds Resolution: SRTM30_PLUS. Marine Geodesy 32, 355-371.
Corliss, J.B., Dymond, J., Gordon, L.I., Edmond, J.M., von Herzen, R.P., Ballard, R.D., Green, K., Williams, D., Bainbridge, A., Crane, K., van Andel, T.H., 1979. Submarine thermal springs on the Galapagos Rift. Science 203, 1073-1083.
Harris, P.T., MacMillan-Lawler, M., Rupp, J., Baker, E.K., 2014. Geomorphology of the oceans. Marine Geology 352, 4-24.
IHO, 2008. Standardization of Undersea Feature Names: Guidelines Proposal form Terminology, 4th ed. International Hydrographic Organisation and Intergovernmental Oceanographic Commission, Monaco, p. 32.
Macdonald, K.S., Scheirer, D.S., Carbotte, S.M., 1991. Mid-ocean ridges: discontinuities, segments and giant cracks. Science 253, 986-994.
Muller, R.D., Roest, W.R., Royer, J.Y., Gahagan, L.M., Sclater, J.G., 1997. Digital Isochrons of the World’s Ocean Floor. Journal of Geophysical Research 102, 3211-3214.
Tunnicliffe, V., Fowler, M.R., 1996. Influence of sea-floor spreading on the global hydrothermal vent fauna. Nature 379, 531 – 533.
Van Dover, C., 2000. The ecology of deep-sea hydrothermal vents. Princeton University Press, Princeton, New Jersey.